|
Kaufman, A.J.1; Olcott, A.2; Sessions, A.3; Brody, K.B.1; Eigenbrode, J.4; Cody, G.4; Geboy, N.1; Williams, B.1; Corsetti, F.A.2; Oliviera, T.F.5; Azmy, K.6; Misi, A.7
1. Department of Geology, University of Maryland, College Park, MD 20742 - kaufman@geol.umd.edu, kbbrody@geol.umd.edu, ngeboy@geol.umd.edu, bwilliams@wam.umd.edu 2. Department of Earth Sciences, University of Southern California, Los Angeles, CA 90089, USA - olcott@usc.edu, fcorsett@usc.edu 3. Division of Geological and Planetary Sciences, California Institution of Technology, Pasadena, CA 91125, USA, - sessions@its.caltech.edu 4. Geophysical Laboratories, Carnegie Institution of Washington, Washington, DC 20015, USA - j.eigenbrode@gl.ciw.edu, g.cody@gl.ciw.edu 5. Companhia Mineira de Metais (CMM), Caixa Postal 03, 38780-000, Vazante, MG, Brazil - flavio@vz.cmm.com.br 6. Department of Earth Sciences, Memorial University of Newfoundland, St. John’s, Newfoundland, Canada A1B 3X5 - kazmy@mun.ca 7. Instituto de Geociencias, Universidade Federal da Bahia, 40170-290 Salvador, Bahia, Brazil - misi@ufba.br
Abstract
Keywords: biomarker, isotope, São Francisco Craton, Neoproterozoic, glaciation
Introduction The Neoproterozoic Bambuí and Vazante groups atop the São Francisco Craton in Brazil are sister basins to the Otavi Group in Namibia on the conjugate Congo Craton, where the ‘snowball Earth’ hypothesis (Kirschvink, 1992) was resurrected. The well-studied Otavi Group hosts at least two cap carbonates atop glacial sediments (Kaufman et al., 1991; Hoffmann and Prave, 1996; Hoffman et al., 1998), but these carbonate accumulations are generally oxidized, lean in organic matter, and have thus not been the focus of biomarker research. In contrast, the conjugate basins where the Bambuí and Vazante groups were deposited in Brazil contain thick intervals of organic-rich shale, limestone, and dolomite interbedded with recognized glacial diamictites. These little metamorphosed sediments are arguably the world’s most organic- and phosphate rich of all Neoproterozoic glacial intervals, are preserved in a basin-wide distribution of well-documented, non-mineralized exploration cores, and provide a unique laboratory for biomarker research. Insofar as biological regulation of greenhouse gases (i.e. CO2 and CH4) at a time of lower solar luminosity is thought to have initiated the Neoproterozoic ice ages (Kaufman et al., 1997; Hoffman et al., 1998; Schrag et al., 2002; Pavlov et al., 2003), biomarker and paleontologic studies of organic-rich pre-glacial shale and carbonate should help to elucidate the source of these gases by identifying the suite of micro-organisms responsible for their buildup. Similar analysis of like lithologies deposited during post-glacial transgression should also explain whether the anomalous accumulations of unusually textured cap carbonates are: 1) the result of silicate weathering under a 10% CO2 atmosphere (Hoffman et al., 1998; Hoffman and Schrag, 2002), 2) a catastrophic methane release from permafrost and methane cold seeps (Kennedy et al., 2001), or 3) ice age bacterial sulfate reduction in the water column similar to the present day #000066 Sea (Gorjan et al., 2000), coupled with extreme rates of primary productivity in the aftermath.
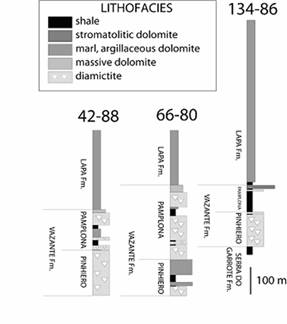
Lithologic and isotopic studies To test the productivity and other ice age hypotheses in well-preserved organic-rich strata, we have conducted biomarker and isotopic studies of shale intervals from the Vazante Group and Lapa Formation. Several cores drilled near Paracatu by GEOSOL and Companhia Mineira de Metais were provided for this study (Figure 1) and were sampled at high stratigraphic resolution through four discrete intervals of organic-rich shale and carbonate.
Figure 1. Lithostratigraphic columns from Morro do Agudo drillholes MAF 42-88, MAF 134-86, and MAF 66-80 of the Neoproterozoic Vazante Group and overlying Lapa Formation in Brazil.
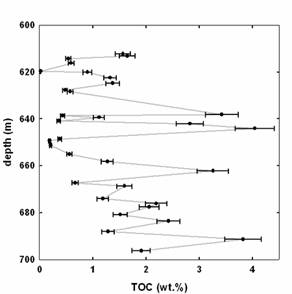
Total organic carbon (TOC) abundances of samples from the Pamplona member (Mocambo) and Lapa Formation shale interval are highly variable, but indicate contents as high as 4.0 (Figure 2) and 1.7 wt.%, respectively, in the studied drillholes.
Figure 2. Total organic carbon (TOC) abundances of shale and marl in the Mocambo shale from drillhole MAF 134-86.
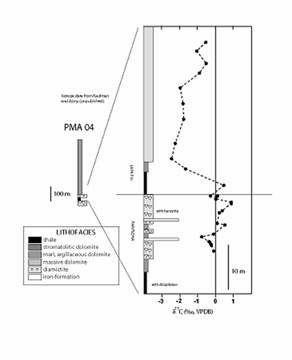
In the Paracatu region, the Pamplona Formation includes diamictite, rhythmic marl, carbonate, and finely laminated, organic-rich #000066 shale, deposited below storm wave base; strata assigned to the Vazante Formation in other regions are primarily composed of carbonate rocks, and are likely either pre- or post-glacial (e.g., Azmy et al., 2001). Evidence for glacial processes is found throughout the Vazante Formation in the studied drillholes. Lonestones, interpreted as glacial dropstones (ice rafted debris), are present throughout the finely laminated shale and indicate the presence of overlying or proximal ice at the time of deposition. Shale samples also contain small nodules with squared prismatic crystal terminations; these formed before significant compaction and are identified as glendonite. Glendonite is a pseudomorph after ikaite, a carbonate mineral that most commonly forms between -1.9 and 7 °C in organic-rich muds, and has been previously reported from other Neoproterozoic glacial successions containing organic matter (Halverson et al., 2004). The stratigraphic architecture and lithologic associations recorded in a field of correlated well logs show that the bituminous shale of the Lapa Formation was deposited during maximal flooding atop a widespread sequence boundary on the upper Pamplona Formation. This erosional surface is overlain by dropstone-laden diamictite and local accumulations of iron-formation, as well as finely laminated carbonate, siltstone and > 10 m of organic- and pyrite-rich shale, which accumulated in the zone of maximum flooding. The stratigraphic succession is consistent with deposition of these lithofacies during post-glacial transgression. This interpretation also fits with carbon isotope trend across the basal Lapa Formation, which reveal a strong negative d13C anomaly (Figure 3).
Figure 3. Lithologic and carbon isotope trends across the upper Vazante Group and lowermost Lapa Formation recorded in drillhole PMA 04.
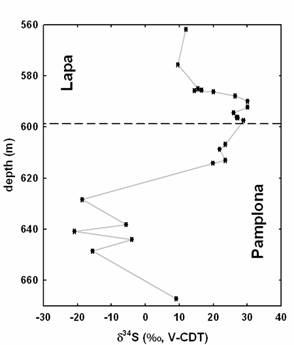
In addition, sulfur isotope analyses of finely disseminated sulfides in the Lapa shale reveal remarkable 34S enrichments (up to +30‰) in basal beds of the formation (Fig. 4). Similar enrichment in both sulfides and trace sulfate in carbonate have been noted in cap carbonate lithofacies worldwide, including successions from Namibia, Death Valley, and from the broadly equivalent Bambuí Group near Belo Horizonte (Hurtgen et al., 2002; Varni et al., 2001). Notably, d34S values in the Lapa Formation decline up section by as much as 20‰. The extremely positive sulfur isotope compositions of these sulfides are likely to be associated with high rates of bacterial sulfate reduction in an anoxic deep glacial ocean with low sulfate concentrations. In this model 32S enriched sulfide would be removed from the ocean through the precipitation of pyrite, leaving residual sulfate in the ocean low in concentration and enriched in 34S. Notably, bacterial sulfate reduction may have also produced the 13C-depleted alkalinity necessary for the ubiquitous cap carbonates that formed rapidly during post-glacial transgression.
Figure 4. Sulfur isotope variations of disseminated sulfide extracted from non-mineralized shale, marl, and diamictite of the upper Pamplona and Lapa formations from drillhole MAF 134-86.
In the aftermath of global glaciation any sulfate or authigenic sulfide of cap carbonate lithofacies should reflect the 34S enrichment (Gorjan et al., 2000; Varni et al., 2001; Hurtgen et al., 2002). The photosynthetic production of oxygen would have driven sulfate reducers back into the sediments in shallow environments. Due to the diffusive limitation of sulfate into sediments, these heterotrophic bacteria would have been cut off from oceanic sulfate and d34S of newly formed sulfides could have risen to values higher than contemporaneous seawater by closed system fractionation. One possible explanation for the up section trend recorded in the Lapa Formation is that sulfate concentrations increased along with productivity, thereby attenuating magnitudes of sulfur isotope fractionation.
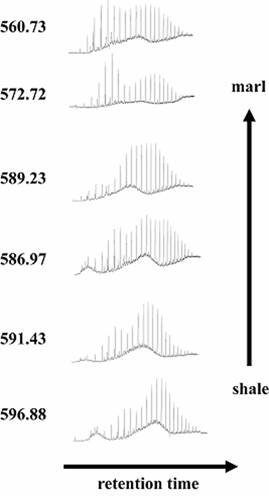
Biomarker studies To further evaluate the possible biological component of the depositional environment of these organic-rich sediments, biomarkers were extracted from the Pamplona and Lapa formation shales by standard techniques (Logan et al., 1999; Brocks et al., 1999; 2003). While these rocks are well preserved in comparison to other syn-and post-glacial deposits of similar age, they are still thermally mature and thus contain small quantities of bitumen. As such, they are especially susceptible to contamination by migrating oil-bearing fluids, drill-core collection and handling, and extant biota. Several lines of evidence support our interpretation that extractable biomarkers in these rocks are autochthonous. In particular, the abundance of taxa-specific biomarkers was higher in the #000066 shale than the marl and carbonate-matrix diamictite, denoting an expected lithological preservation bias (Fig. 5). GC-MS-MS analyses of biomarkers extracted from the syn-glacial organic-rich shales include 2-a methylhopanes (indicative of cyanobacteria; Summons et al., 1999) and trimethyl arylisoprenoids (green and purple sulfur bacteria; Koopmans et al., 1996). These biomarkers provide firm evidence for the presence of photosynthetic organisms, and by extension the penetration of sunlight to the sea surface. The green sulfur bacteria are anaerobic organisms that utilize H2S as the electron donor for photosynthesis, and are strongly inhibited by the presence of O2. Their presence in other sediments is widely interpreted as evidence for an anoxic, sulfidic photic zone. This anoxic water would allow for the accumulation and subsequent precipitation of iron, a phenomenon observed in glacial assemblages (Kaufman et al., 1991; Kirschvink, 1992). Other extracted biomarkers include 3-b methylhopanes, known only from microaerophilic proteobacteria including the aerobic methanotrophs (Summons and Jahnke, 1992). Gammacerane, a C30 triterpane, is derived from protozoan tetrahymanol, and while the organisms that produce tetrahymanol are heterotrophic aerobes, gammacerane is often prevalent in sediments formed under a stratified water column. Stable carbon isotopic studies of gammacerane have shown that the bacteriovores that produce tetrahymanol consume green and purple sulfur bacteria, so the protozoa live at or below the chemocline (Sinninghe Damste et al., 1995). C27 and C28 di- and trinorhopanes tend to be found in high concentration in sulfidic sediments, although the precursor organism is not yet known. Steranes that are not methylated at C-4 are produced exclusively by aerobic eukaryotes. In sum, the biomarker data from the São Francisco Craton describe a diverse microbial community living in a stratified water column based upon photosynthetic primary production, consistent with significant sunlight availability and thus with relatively thin sea ice or an ice-free tropical ocean. This view is inconsistent with the depauperate record of both microfossils and biomarkers both before and after the Neoproterozoic ice ages (Corsetti et al., 2003), as well as the biotic crisis suggested by the ‘snowball Earth’ hypothesis.
Figure 5. Total ion chromatograms of bitumen from six Lapa Formation samples in drillhole MAF 134-86.
References Azmy, K., Veizer, J. Misi, A., de Oliveira,
T. F., Sanches, A.L., Dardenne, M.A. 2001. Dolomitization and isotope stratigraphy of the Vazante
Formation, São Francisco Basin, Brazil. Precambrian Research, v. 112:303-329. Brocks, J.J., Logan, G.A.,
Buick, R., Summons, R.E. 1999. Archean molecular fossils and the early
rise of eukaryotes. Science, v. 285:1033-1036. Brocks,
J.J., Buick,
R., Logan,
G.A., Summons,
R.E.
2003. Composition and syngeneity of molecular fossils from the 2.78
to 2.45 billion-year-old Mount Bruce Supergroup, Pilbara Craton, Western
Australia. Geochimica et Cosmochimica Acta, v. 67:4289-4319. Corsetti,
F.A., Awramik, S.M., Pierce, D. 2003. A complex microbiota from snowball
Earth times: Microfossils from the Neoproterozoic Kingston Peak Formation,
Death Valley , USA. Proceedings of the National Academy of Sciences,
v. 100:4399-4404. Gorjan, P., Veevers, J.J., Walter, M.R. 2000. Neoproterozoic sulfur-isotope variation in Australia and global implications. Precambrian Research, v. 100:151-179. Halverson, G.P., Maloof, A.C.,
Hoffman, P.F. 2004. The Marinoan glaciation (Neoproterozoic) in northeast
Svalbard.
Basin Research, v. 16, 297-324. Hoffman, P.F. & Schrag,
D.P. 2002. The snowball Earth hypothesis: testing the limits of global
change. Terra Nova, v. 14:129-155. Hoffman, P.F., Kaufman, A.J.,
Halverson, G.P., Schrag, D.P. 1998. A Neoproterozoic snowball Earth.
Science, v. 281:1342-1346. Hoffmann, K.-H. & Prave,
A.R. 1996. A preliminary note on a revised subdivision and regional
correlation of the Otavi Group based on glaciogenic diamictites and
associated cap dolostones. Geological Survey of Namibia Communications,
v. 11:77-82. Hurtgen, M.T., Arthur, M.A.,
Suits, N.S., Kaufman, A.J. 2002. The sulfur
isotopic composition of Neoproterozoic seawater sulfate: implications
for a snowball Earth? Earth and Planetary Science Letters,
v. 203:413-429. Kaufman, A.J., Hayes, J.M.,
Knoll, A.H., Germs, G.J.B. 1991. Isotopic compositions of carbonates
and organic carbon from Upper Proterozoic successions in Namibia:
Stratigraphic variation and the effects of diagenesis and metamorphism.
Precambrian Research, v. 49:301-327. Kaufman, A.J., Knoll, A.H.,
Narbonne, G.M. 1997. Isotopes, ice ages, and terminal Proterozoic
Earth history. Proceedings of the National Academy of Sciences, v.
94:6600-6605. Kennedy, M.J., Christie-Blick,
N., Sohl, L.E. 2001. Are Proterozoic cap carbonates and isotopic excursions
a record of gas hydrate destabilization following Earth’s coldest
intervals. Geology, v. 29:443-446. Kirshvink, J.L. 1992. Late
Proterozoic low-latitude global glaciation: the snowball Earth. In
The Proterozoic Biosphere. In: Schopf, J.W. & Klein, C.(Eds.)
Cambridge University Press, Cambridge:51-52. Koopmans, M.P., Schouten, S.,
Kohnen, M.E.L., Sinninghe Damste, J.S. 1996. Restricted
utility of aryl isoprenoids as indicators for photic zone anoxia
Geochimica et Cosmochimica Acta, v. 60:4467. Logan, G. A., Calver, C.R., Gorjan, P., Summons,
R.E., Hayes, J.M., Walter, M.R. 1999. Terminal Proterozoic mid-shelf
benthic microbial mats in the Centralian Superbasin and their environmental
significance. Geochimica et Cosmochimica Acta, v. 63: 1345-1358. Olcott, A.N., Sessions, A.L.,
Corsetti, F.A., Kaufman, A.J., Oliviera, T.F. (in review) Direct
evidence for significant primary production during widespread Neoproterozoic
glaciation. Science. Pavlov, A.A., Hurtgen, M.T.,
Kasting, J.F., Arthur, M.A. 2003. Methane-rich Proterozoic atmosphere?
Geology, v. 31:87-90. Schrag, D.P., Berner, R.A.,
Hoffman, P.F., Halverson, G.P. 2002. On the initiation of a snowball
Earth. Geochemistry, Geophysics, Geosystems, v. 3, no. 6. Summons, R.E., Jahnke, L.L.,
Hope, J.M., Logan, G.A. 1999. 2-Methylhopanoids as biomarkers for
cyanobacterial oxygenic photosynthesis. Nature, v. 400:554. Summons, R.E. & Jahnke,
L.L. 1992. Hopenes and hopanes methylated in ring-A; correlation of
the hopanods from extant metylotrophic bacteria with their fossil
analogues. In: Moldowon, J.M., Albrecht, P., Philp, R.P., Prentice,
H. (Eds.) Biological markers in sediments and petroleum; a tribute
to Wolfgang K. Seifert (Englewood Cliffs, NJ, 1992). Sinninghe Damste, Jaap S.,
van Duin, A.C. T., Hollander,
D., Kohnen,
M.E.L., de
Leeuw, J.W.
1995. Early
diagenesis of bacteriohopanepolyol derivatives; formation of fossil
homohopanoids Geochimica et Cosmochimica Acta, v. 59:1895. Varni, M. A., Kaufman, A.J.,
Misi, A., Brito Neves, B.B. 2001. Anomalous d34S
signatures in trace sulfate from a potential cap carbonate in the
Neoproterozoic Bambui Group, Brazil. Geological Society of America
Abstracts with Programs, v. 33:96.
|
||||||||||||||||||||||||